The essence of the immune system—a force of our vitality that hitherto had been taken for granted—gained worldwide attention with emergence of HIV-AIDS in the 1980s. Our immune system is the molecules, cells, and tissues utilised by the body to protect itself from foreign substance, especially invading microorganisms, but also tissue injury and preventing malignancy. Immunity is the state of protection from, or resistance to, infection — a consequence of a prior exposure to an infection or a vaccine, or the passive transfer of antibodies. Immunodeficiency or hypersensitivity (including autoimmunity) are the hallmarks of immunopathology.
The Immune System
The immune system has the inherent capacity to rapidly respond to any pathogen through a non-specific innate clonal expansion, without the need to await an antigenic response, thereby preventing tissue invasion but at the same time triggering the more specific adaptive immune response which can be recruited at a later time, if and when required. Innate and adaptive immunity are distinct—but interdependent—entities.
Physical barriers to pathogen entry—such as epidermal intercellular tight junctions, coughing, sneezing, the acidic environment of the stomach, alimentary tract enzymes, the flushing effect of urinary flow and urination, the resident microbial flora—can, more broadly speaking, also be considered part of the innate immune response.
But once a pathogen manages to breach an epithelial barrier, it will immediately encounter cells with receptors that recognise its surface antigens, usually phagocytic macrophages. Engulfing the pathogen, the macrophage releases chemical messengers (cytokine) that signal to other cells to modify their behaviour as well as chemicals (chemokine) that attract other cells to their location—the site of infection.
There are also plasma proteins that bind to bacterial cell surface receptors. Once activated, these complement proteins attract phagocytes to come and attack the bacteria—thereby also acting as chemo-attractant.
Acting together, these aspects of innate immunity lead also to dilation of blood vessels and increased permeability of blood vessel walls, and neutrophil recruitment. Very quickly, heat, redness, pain, and swelling can be discerned in the affected tissue—the cardinal signs of inflammation. Neutrophils are the predominant bacterial (and fungal) phagocyte, assisted by macrophages — together known as acute inflammatory cells. They form the first-line of defence against invading pathogens (and also for presenting to lymphocytes the responsible antigen). They are also key to subsequent tissue repair.
Only later are lymphocytes recruited to this site of inflammation, after being called upon by the antigen-bearing dendritic cells (surveillance drones of the immune system that ingest antigen and other foreign extracellular material, including virus-infected host cells, by macro-pinocytosis) that have since migrated to meet them at the local draining lymph node. As the lymphocytes make their way to the inflammatory focus, they also recruit further neutrophils and macrophages. Gradually, in evolving inflammation, lymphocytes come to predominate over neutrophils (and macrophages), at which point the inflammation is designated chronic.
Antigen is a compound word made up of the first parts of the words “antibody generation”, but is now accepted to refer to any substance that can induce an adaptive immune response (irrespective of antibody generation).
Tissues and organs
All blood cells originate in the bone marrow as derivatives of one pluripotent haematopoietic stem cell, which divides along two major cell lines — lymphoid and myeloid progenitors.

Cells of the immune system are transported through specialised lymphatic vessels (as well as in the bloodstream). The lymphatic vessels are one-way valved channels interspersed by small glands of lymphocytes, the draining lymph nodes.

There are other, albeit less discreet, areas of concentrated lymphoid tissue. They share a common micro-architecture, designed to trap the antigen that is on the presenting cell and initiate adaptive immunity. These are:
- MALT (mucosa-associated lymphoid tissue), such as the Waldeyer’s tonsillar ring, is a diffuse system of small concentrations of lymphoid tissue which play a role in regulating mucosal immunity. It has several components:
- GALT — gut-associated lymphoid tissue, e.g., Peyer’s patches
- BALT — bronchus-associated lymphoid tissue
- spleen
Lymphoid
There are two primary lymphoid organs — the bone marrow, and the thymus gland. The lymphoid progenitor, in turn, differentiates along one of two lines — lymphocyte and dendritic cells.
The lymphocytes are classed into 3 types:
- T cells
- B cells
- NK (Natural Killer) Cells
B cells originate and mature in the bone marrow. T cells originate in the bone marrow but migrate to the thymus for maturation. Lymphocytes are maintained and presented with antigens for initiation of adaptive immunity in secondary lymphoid organs. There is a B cell with a receptor for virtually every foreign antigen. The receptor itself is the antibody that will later be secreted in large quantities upon stimulation of that cell. Once stimulated, B lymphocytes differentiate into these antibody-secreting plasma cells.
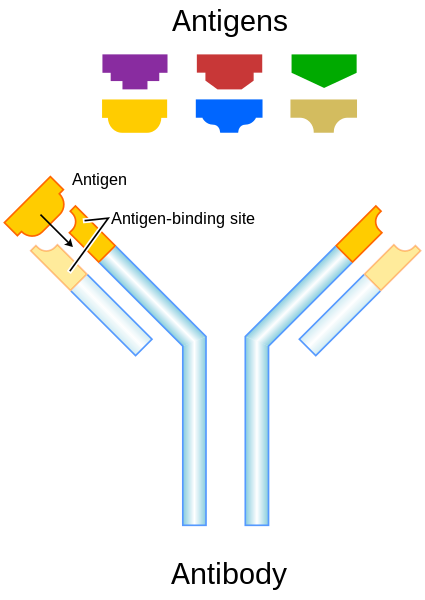
Differentiated B lymphocytes, known as plasma cells, secrete proteins that recognises a specific antigen. These antibodies have s structure that includes a constant region of limited type, and helping to dispose of the antigen once bound, and an antigen-specific variable region, which combines directly with the antigen. Neutralisation occurs when the antibody binds the pathogen antigen, blocking the pathogens entry into the cell and also creating—at the Ab-Ag interface—a receptor for the first component of complement (C1). This activates a complement cascade of reactions whereby the ultimate proteins attack and destroy the pathogen’s cell wall. Phagocytes then recognise the constant region of the antibody and proceed to destroyed the antigen, known as opsonisation. Adaptive immunity involving B cells and antibodies is known as humoral immunity.

{kind=link}
There are two main types of T cells. One type of T cell, recognised by its surface “CD4” receptors, directs the other immune cells and is designated the T Helper or CD4 cell. The other T-cell type (recognised by its surface “CD8” receptors), once stimulated, kills virus-infected cells and is termed a T Suppressor or CD8 cell. T lymphocytes also contain surface receptors for virtually every conceivable foreign antigen. Adaptive immunity involving T cells and their cytokines is known as cellular immunity.
It is during the maturation process in the bone marrow (B cells) and thymus (T cells) that naïve lymphocytes come to bear receptors with just one single specificity, though millions of different recombinations in the genes that encode for their surface receptors. And any lymphocytes created that are potentially self-reacting, undergo clonal deletion during development.
Clonal expansion in adaptive immunity: 1 becomes 1000
Upon being presented with its specific antigen (by an antigen-presenting cell or APC) a naïve circulating lymphocyte (B cell or T cell) will stop circulating and grow in size to become a lymphoblast, beginning a phase of rapid division into perhaps a thousand identical daughter cells. Activated lymphocytes also express cell surface adhesion molecules which allow them to migrate into or remain in the site of infection.
It is not enough that the lymphocyte receptor bind with its antigen for lymphocyte activation. A second signal—a co-stimulatory molecule—is needed. The antigen presenting cell (APC) provides this second signal for the T cell. The T cell provides this second signal for the B cell.
Pathogens that are located within a host cell—viruses, some bacteria and other pathogens—are out of reach of antibodies. These host cells do, however, display pathogen antigens on their surface. T cells (CD8) detect these antigens and destroy the cell. Some pathogens located within macrophages may be incorporated into lysosomes within the macrophage, which contain enzymes that destroy the pathogen. Others—Mycobacterium tuberculosis for instance—may be contained within, and replicate in, storage vesicles that do not fuse with lysosomes and therefore survive.
A TH1 subset of CD4 T-cells have the ability to induce fusion of the lysosome with the pathogen-containing vesicle by activating these macrophages and other bactericidal mechanisms (as well as further chemo-attractants for recruitment of further macrophages). Another subset of CD4 T-cells—TH2 cells— stimulate B cells to produce immunoglobulin (antibody) which can destroy extracellular antigen. They also stimulate B cells to produce the allergic type of antibodies responsible for allergic diseases.
Most effector lymphocytes undergo apoptosis (programmed cell death) once infection has subsided. Some remain as memory cells. Memory cells bestow upon the immune system early-recognition capabilities for mounting an effective and specific immune response sooner. Immunologic memory is the basis for immunity, and immunisation.
Natural killer cells are a less common T-cell type that survey the tissues for any abnormality—such as tumour cells or cells infected by virus—and non-specifically (i.e. do not display antigen-specific receptors) eliminate these cells.
The Major Histocompatibility Complex (MHC)
Glycoprotein molecules that capture foreign antigen within the cell and display fragments of these foreign antigens on the cell surface for interaction with T lymphocytes are encoded by a cluster of genes known as the MHC. The MHC molecule contains a cleft where the fragment of foreign antigen is held and taken to the cell surface. T cells then recognise the specific MHC molecule-antigen combination.
- MHC-Class I molecule
- combine with virus particles that are produced in the cell cytoplasm and display the viral antigens on the cell surface
- recognised by CD8 T cells
- MHC-Class II molecule
- combine with antigen fragments that are contained within vesicles within cells; this includes antigen bound by antibody after the macrophage has engulfed it
- recognised by CD4 T cells
T-cells must express both the antigen receptor, and a co-receptor that recognises MHC-I (CD8) or MHC-II (CD4). Once stimulated by combination with the target antigen, T cells release a large array of cytokines to allow them to fulfil their effector function.

Myeloid
The myeloid progenitor differentiates into a number of different blood cell types, including neutrophils, macrophages, dendritic cells, eosinophils and basophils, as well as erythrocytes and platelets.
The Cells
- B lymphocytes
- T lymphocytes
- Neutrophils
- Macrophages
- Dendritic cells
- NK cells
The Situation
- Inflammation
- Immunity
The Rules
Manipulation of the immune system to effect or modify immunity, inflammation, or tissue repair in:
- Allergy
- Autoimmune disease
- Infection
- Malignancy
- Transplantation
References
- Basic Immunology: Brad Frankum. Professor of Clinical Education, UWS.
- Mucosa-associated lymphoid tissue. Wikipedia. Available at https://en.wikipedia.org/wiki/Mucosa-associated_lymphoid_tissue as at 10 July 2016.